虾类用什么呼吸(动物呼吸腮到书肺的过渡)
一、呼吸内脏团的出现
(一)皮毛呼吸到呼吸腮

图1-3-1 软体动物腮呼吸结构示意图
软体动物是由环节动物演化而来,朝着不很活动的生活方式较早分化出来的一支。身体柔软,大多数具有贝壳。身体柔软,一般左右对称,不分节可分为头、足、内脏团三个部分,体被外套膜,常常分泌有贝壳。次生体腔,后肾管,螺旋式卵裂,个体发育中具有担轮幼虫。鳃或肺呼吸。
软体动物身体背部皮肤皱褶向腹面延伸面形成的一种保护器官,由内、外表皮、中间的结缔组织和少数肌肉纤维组成。外套膜与内脏之间有外套腔,外套腔内有鳃和口、肛门、肾脏、生殖腺的开口。外套膜的边缘构造很复杂,常具各种形状的触手,有的种类有外套眼。外套膜内表皮细胞具纤毛,纤毛摆动,造成水流,使水循环于外套腔内,借以完成呼吸、排泄、摄食等。
软体动物体表呼吸几乎完全退化,发育出围心腔、生殖腺内腔、排泄器内腔三个腔体,呼吸器官“腮”结构与心脏和肾脏分布于三个腔体内,有脉管相连,间介中胚层和脏壁中胚层已经分化发育,说明三个胚层已经在体腔内汇集,皮肤呼吸功能基本退化完毕,皮肤神经迁移集中于腹部形成腹索中枢神经结构。(见图1-3-1)
(二)皮腮-呼吸树

图1-3-2过皮鳃的切面示意图

图1-3-3 呼吸树结构示意图
棘皮动物门是一类后口动物,在无脊椎动物中进化地位很高。它们在形态结构与发生上都有一些独特之处,与原口动物有很大不同。外观差别很大,有星状、球状、圆筒状和花状。成体五放辐射对称,由管足排列表现出来。身体区分为有管足的辐部或步带和无管足的间辐部或间步带。内部器官,包括水管系、神经系、血系和生殖系均为辐射对称,只有消化道除外。身体有口面和反口面之分。骨骼很发达,由许多分开的碳酸钙骨板构成,各板均由一单晶的方解石组成。多为雌雄异体,生殖细胞释放到海水中受精,幼体在初发生时形状相同,以后则随纲而异,少数种类可行无性裂体繁殖。对水质污染很敏感,再生力一般很强。摄食方式为吞食性、滤食性和肉食性。
海星和蛇尾的消化系有一大囊状胃,肠直走,不弯曲;海星常有从胃向各腕伸出的一对称为幽门盲囊的消化腺。其他各纲的消化管多为一长而弯曲的管子,由肠系膜连于体壁。蛇尾和某些海星无肛门,凡是不能消化的东西从口里吐出。海百合的口和肛门全朝上,许多歪形海胆肛门常在体后部或口面。没有明显的排泄器官,排泄功能由变形游走细胞负责。呼吸系也不发达。海星的皮鳃(见图1-3-2)和海胆的围口鳃可增加呼吸的能力和面积。某些海参有特殊的呼吸器官,称呼吸树(见图1-3-3)。
棘皮动物门呼吸系统出现两种呼吸结构,皮腮结构时原始皮毛呼吸演变的产物,而呼吸树的出现意义很大,说明内胚层肺分化支已经开始分化发育。
动物从半消化管到完整消化管演变过程中,出现三个内胚层分化支,首先是随着消化管后移分化发育外胚层内移形成的内胚层肺分化支;第二是原肠后端向体腔内分化的内胚层肝脏分化支;第三是排泄孔内陷形成的内胚层膀胱分化支。到棘皮动物门虽然内脏器官还都是内脏团,但是三条内胚层分化支发育出了内脏器官的原基结构,说明已经在向内脏系统演化。
虽然棘皮动物门还有皮表呼吸的皮腮器官存在,呼吸树结构非常类似高等动物的肺结构,这种体内呼吸器官的出现意味着呼吸功能已经内迁到体内,意味着皮表呼吸退化就要结束了。
(三)气管-书肺

图1-3-4 气管熟肺结构示意图
节肢动物门,是动物界最大的一门,通称节肢动物,包括人们熟知的虾、蟹、蜘蛛、蚊、蝇、蜈蚣以及已绝灭的三叶虫等,身体两侧对称。由一列体节构成,异律分节,可分为头、胸、腹三部分,或头部与胸部愈合为头胸部,或胸部与腹部愈合为躯干部。例如:昆虫纲(蝗虫)动物身体分头、胸、腹三部分;甲壳纲(虾)动物身体分头胸、腹二部分;蛛形纲(蜘蛛)动物身体分头胸部、腹部;多足纲(蜈蚣)动物身体分头部、躯干部。身体的分部在生理机能上也出现了分工:头部:感觉和取食中心;胸部:运动和支持中心;腹部:营养和繁殖中心。
节肢动物的体腔在发育早期也形成中胚层的体腔囊,但在继续发育的过程中,不扩展为广阔的真体腔,而是退化为生殖管腔、排泄管腔和围心腔。在以后的发育过程中,围心腔壁消失,使体壁和消化道之间的初生体腔与围心腔的次生体腔相混合,形成混合体腔。混合体腔内充满血液,混合体腔也称作血腔。
⑴有爪纲也称原气管纲,如栉蚕等。
⑵肢口纲体分头脑部和腹部。头脑部有6对附胶,即一对螫肢和5对步足;无触角。腹肢7对。用鳃呼吸。如鲎等。
⑶蛛形纲体分头胸部和腹部。头胸部有6对附肢,即一对赘肢、一对脚须(触肢)和4对步足;也无触角。腹肢几乎完全退化。用书肺和气管呼吸。如各种蜘蛛等。
⑷甲壳纲体常分头脑部和腹部。头胸部有13对附肢,即5对头肢和8对胸肢。5对头肢包括2对触角,一对大颚和2对小颚。8对胸肢中前几对为颚足,其余为步足。腹肢有或无。用鳃呼吸。如各种虾和蟹等。
⑸多足纲体分头部和躯干部。头部有3一4对附肢,即一对触角、一对大颚,和1~2对小颚。躯干部有多对步足,每一体节l一2对。用气管呼吸。如蜈蚣等。
⑹昆虫纲体分头、胸、腹三部、头部有4对附肢,包括一对触角、一对大颚,一对小颚以及一对左右愈合成为一片的下唇、胸部有3对步足。腹部附肢几乎完全退化。如各种蚊和蝇等。
可见节肢动物的呼吸器官,因适应各种环境,可由各种变化。水生类型用腮或书肺呼吸。陆生类型用类似书肺但内陷并有小孔与外界相通的书肺或用分枝状的气管呼吸。有些类型用体面直接呼吸。
二、呼吸系统发育发生机制
(一)卵裂方式与动物进化
三胚层动物内胚层分化发育,从半消化腔到完整消化腔演化过程中,内胚层原肠胚分化出向体腔延伸的三条分化支:肺分化支,肝分化支,膀胱分化支。三条内胚层分化支从原肠外侧前中后三个位置向体腔内衍伸分化发育,与中胚层耦合发育出现了不平衡性,从软体动物门到节肢动物,动物的呼吸组织器官分布出现明显的多样性,软体动物门的呼吸腮位于肛门附近;棘皮动物的皮腮位于体表,呼吸树在肛门两侧;节肢动物的气管,气囊遍布全身,疣足动物的书肺位于腹部,器官开孔于疣足。为什么动物呼吸器官的演化出现如此多样性呢?根据现代动物发育发生学分析,这一现象与不同卵裂方式相关。
卵子受精以后,受精卵完成父母本遗传物质的融合和细胞质重排,便开始了多细胞有机体的形成过程。这一过程开始于卵裂。从受精向卵裂的转变是由促有丝分裂因子活化而实现的。受精卵经过多次重复的有丝分裂形成很多细胞的过程称为卵裂。卵裂所形成的细胞称为卵裂球或分裂球。卵裂类型基本分两种:完全卵裂,不完全卵裂。
完全卵裂:
分裂面将分裂球完全分割成两半,子细胞完全分开。多数动物的卵,不仅卵黄分布均匀,分出的分裂球的大小一般相差不多,是为全裂;有些卵黄分布不均匀的卵,如两栖类和一些低等鱼类的卵,也是全裂。但第3次分出的裂球大小差异较大,在动物极的体积小,称为小裂球,在植物极的含卵黄多,体积大,称为大裂球。
全裂又可根据分裂球排列的形式,分成6种类型:
1. 辐射型
第3次卵裂以后,上层的分裂球很整齐地排列在下层之上,呈辐射排列,如棘皮动物和腔肠动物等。
2. 螺旋型
第3次卵裂时,纺锤体不是和赤道面垂直而是倾斜呈45°角,分出的动物极的分裂球位于两个植物极分裂球之间,以后的卵裂也是倾斜的,按顺时针方向倾斜的,称右旋型;按逆时针方向倾斜的,称左旋型。往往是右旋、左旋交替排列,如多毛类环节动物,软体动物中的腹足类和瓣鳃类等。
3. 两侧对称型
卵裂的情况和辐射型的相似,主要是在第1次卵裂后,已呈现两侧对称,以后多次卵裂均保持这种形式。幼虫和成体的两侧对称面和第1次分裂面相符,如两栖类和尾索动物等。
4. 两轴对称型
在第2次卵裂后,4个分裂球不仅左右对称,前后也对称,以后的卵裂,始终保持这种情况,因此称为两轴对称,如栉水母。
5. 混合型
最初是螺旋型,以后是对称型,混合进行。如一些环节动物。
6. 不规则型
分裂球的排列和卵裂的程序均是不规则的,如蛔虫的第2次卵裂后分裂球的排列;又如哺乳动物的卵,从第2次卵裂起,分裂球就不规则地排列。
不完全卵裂:
也称为偏裂。大量卵黄集中于卵的一端或卵的中央,以致分裂面不能将卵分成两部分,仅卵质部分被分开,含卵黄的部分仍连在一起。不全裂又可分为两个类型:
1.盘状卵裂卵裂仅在动物极的胚盘上进行,胚盘下的大量卵黄部分不分裂,如软体动物中的头足类,软骨鱼、硬骨鱼、爬行类和鸟类。
2.表面卵裂卵黄集中于卵的中央,最初几次卵裂只是细胞核在卵黄里的原生质岛中进行,细胞核达到一定数目、进入卵黄四周的卵质中,在表面继续进行分裂,如昆虫。
动物愈低等,卵子的构造也愈原始和简单,一般都是全裂,分裂球大致相等。最低等的多细胞动物,如海绵动物、腔肠动物等是辐射型对称体型,扁形动物以上的动物都是两侧对称的体型。卵裂反映了这种基本情况。
脊索动物的卵裂比较明显地反映出进化的过程。原始的半索动物有的是螺旋型卵裂,一般是辐射型卵裂。尾索动物(海鞘类)和头索动物(文昌鱼)为两侧对称型卵裂。到鱼类,卵黄逐渐增多并集中,相应地从全裂发展到盘状卵裂。从肺鱼到两栖类、爬行类、鸟类和哺乳类,表现为端黄卵的演变过程。肺鱼和两栖类是全裂,但植物极的分裂速度比较慢。爬行类和鸟类是盘状卵裂。最低等的卵胎生哺乳动物(单孔类)仍是盘状卵裂。到高等的胎生哺乳动物,因胚胎的营养物质直接从母体胎盘获得,卵黄大量减少,又恢复全裂,但仍然保留盘状卵裂的痕迹。因此,高等哺乳动物的均黄卵与其他均黄卵不同,是次生均黄卵。
从动物胚胎卵裂的两种方式看出,越原始的动物越偏于完全卵裂法,这是因为三个胚层分化支还没有完全特异化定性,还无法进行特异组织器官发生发育;而到不完全卵裂说明胚层分化的特异性越来越强,特定的分化支发育特定的组织器官,这就是不完全卵裂现象,由此,从完全卵裂到不完全卵裂的比较可以作为动物演化程度判断的一项标准。
(二)原肠胚分化与呼吸器官发生
1. 原肠形成方式与呼吸器官发育
无论哪一种卵裂法分化发育,呼吸消化同源性都是一样的,可以通过原肠胚消化呼吸同源性来找到呼吸器官演化的总体规律。
囊胚继续发育,囊胚的细胞通过细胞迁移,重新排列和再组合,形成双胚层或三个胚层的胚胎,叫做原肠胚。在这一过程中,囊胚的部分细胞通过不同的方式迁移到囊胚内部,形成原肠。留在外面的的细胞组成的胚层称为外胚层,迁移到内部的细胞组成中内胚层。由囊胚形成原肠胚的细胞运动或迁移过程称为原肠形成或原肠作用;也称为形态发生。
原肠形成的方式和过程比较复杂,往往是几种不同的细胞迁移运动先后综合进行的。而且不同的动物其原肠运动也会有所不同。原肠胚形成的细胞迁移运动方式一般有如下几种:(见图1-3-5)
(1)移入:腔囊胚的部分细胞,从外层经过增殖而迁移到囊胚腔中,逐渐将囊胚腔填满。有两种移入方式,一种是从植物极单极移入,另一种是细胞同时从多处相内移入。
(2)分层:囊胚表面的一层细胞分裂形成两层。有的腔囊胚向内分出一层,成为内层(水母)。有的实心囊胚则向外分出一层,成为外层(水螅,水母)。脊椎动物中也有通过分层法形成胚层的。
(3)内陷:植物极的细胞成片地同时向内陷入,并逐渐扩大而形成内外两层。陷入的细胞围成的腔称为原肠腔。原肠的口逐渐缩小为一圆形的孔,称为胚孔。胚孔的边沿按其位置分为背唇、腹唇和两个侧唇。
(4)内卷:在腔囊胚或盘状囊胚中,表面的细胞从胚孔或从胚盘的边沿,经过连续的细胞迁移而向内卷入。鱼类、两栖类、爬行类和鸟类等原肠形成过程中都有这种细胞的运动方式。
(5)外包:鸟类、鱼类盘状囊胚动物极表面的细胞沿着胚盘的边沿向植物级延伸包围。这种方式称为外包。
(6)集中和延伸:在胚胎细胞内卷的同时,囊胚或原肠胚表面的细胞向胚孔或原条处集中。当胚胎出现了头尾轴而伸长时,外部和内部的细胞群也随着胚体而延伸。
在原肠胚形成时,无论是胚胎表面或是进入了内部的细胞都在连续地运动着;方式也不是单一的,一般是几种不同的运动方式随着时间和空间联合进行。而每一个物种原肠胚形成时的细胞运动都是按照规定的模式进行的。

图1-3-5 原肠胚发育运动示意图
2. 胚胎极性与呼吸器官发生
动物卵细胞的富含原生质的一端,由于卵内所含细胞质、细胞器、核糖体、卵黄、色素粒及糖原颗粒等物质的不均匀分布而表现出极性,分为动物极和植物极;营养物质(卵黄)较少、卵裂速度较快的一极称为动物极;细胞核偏位于动物极。与动物极相对的一端含较多的卵黄颗粒或卵黄小板、卵裂速度较慢的一极称植物极。由于卵黄较其它细胞的比重大,植物极总是向下,卵内各种物质往往沿卵轴形成梯度。(图1-3-6)

图1-3-6动植物极发生发育示意图
动物胚胎发育极性的出现成为一种预定模式,内胚层原肠成为胚胎植物极,外胚层成为胚胎动物极,因为植物极总是向下分化,诱导外胚层细胞随之向内迁移,这就是动物胚胎发育的内卷运动机理,形成胚胎发育背--腹轴性;而位于前后前端的中胚层和内胚层诱导神经胚从前后两端内旋迁移耦合,形成胚胎发育的前--后轴性;由于神经胚细胞从胚体中枢神经中轴线位置向远端迁移分化,形成胚体远--近轴性,三种胚胎轴性发育使得动物体具有了空间极性定位定性,呼吸组织结构也呈现出极性定位(见图1-3-7)
(1)背-腹轴性与体表呼吸器官
胚体内卷将植物极内卷成管状模式发育出原肠结构,是胚层动物发生发育的第一阶段模式,动物胚胎模式形成了动物胚胎背—腹轴性,腹侧原肠发育成消化吸收功能的肠管结构,背侧发育出体壁担负呼吸神经功能的皮表结构,原始动物体表呼吸器官发生的原理就是胚胎背-腹轴性形成机制。
(2)前-后轴性与前后呼吸器官
原肠胚发育肠道结构经历从前向后逐渐发育的过程,也就是从半消化管向完整消化管演化的过程,在这一从前向后发育的过程中,后端排泄孔从后向前发育,这样原肠发育出现了前后内旋发育两条分化路线,这一分化运动被胚胎预定成固定模式后,就出现了胚胎前-后轴性。体表就会随着前-后轴内卷向体内,在前-后轴位置都会出现呼吸器官的发生,这就是原始动物呼吸器官前后都有现象发生的机理。
(3)远-近轴性与远近呼吸器官
原肠胚从半消化管向完全消化管过度发育过程中,原肠中央的消化腺分化支从原肠内侧向外侧分化发育,经过中胚层延伸到外胚层时耦合,就会产生内部消化器官和外部消化器官。
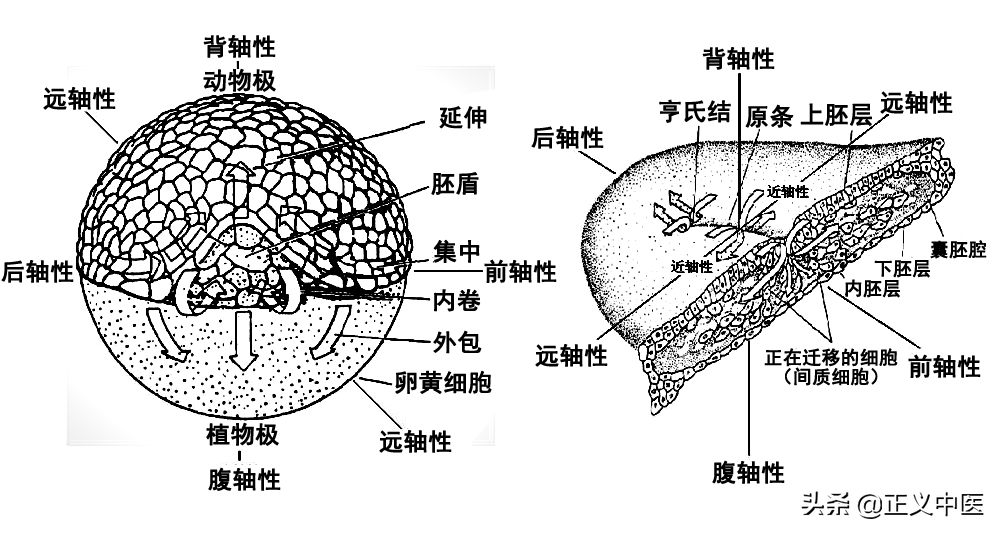
图1-3-7 胚胎发育极性示意图
动物胚胎极性发育分为前后两个演化阶段,前演化阶段叫原口动物,胚胎时期的胚孔(原口)发育成动物的口,肛门是在相对的一侧开口形成的,节肢动物以前的无脊椎动物类群属于原口动物,原口动物的另一个特点是以裂体腔法形成体腔.包括:扁形动物,线形动物,环节动物,软体动物,节肢动物.,这种动物主要是在胚胎背-腹轴性,前-后轴性形成中发育而来,所以呼吸器官的发生分布比较原始。
后演化阶段叫后口动物,在胚胎发育中原肠胚期,其原口形成动物的肛门,而在与原口相对的一端,另形成一新口的动物称为后口动物。后口动物的胚胎发育是辐射不定型卵裂;在原肠期的后期,与原口相反一端的内外两胚层相互贴紧,最后穿成一孔,成为幼虫的口,后口动物因此得名。这些动物的原肠背部两侧,内胚层向外突出成对的囊状突起体腔囊,体腔囊和内胚层脱离后,在内、外胚层之间逐步扩展成为中胚层。这种形成方法称为肠体腔法(见体腔动物)。半索动物门、毛颚动物门、棘皮动物门、脊索动物门动物均为后口动物。它们是动物进化的主干。这种胚胎发育模式使得动物呼吸器官逐渐集约化到前部,说明胚胎远-近轴性发育模式已经基本形成,人体也是这一支系而来。
相关文章
-
练好这三位打野英雄!s36赛季上王者简单!
在王者荣耀S36赛季中,打野的位置还是比较重要的,就是因为打野有着带起前期节奏的重要性,本期咱们推荐的这三位英雄具有较高的上分潜力,适合不同水平的玩家用来冲击更高段位,尤其是目标直指王者印记的玩家。一起来看看都有谁上榜了吧:典韦:经过加强后,典韦的机动性和生存能力得到了提升,一技能的小位移和净化效果
2024-11-23 10:30:18
-
道天录保姆级天元矿脉玩法详解
>>基础介绍<<天元矿脉需要使用矿锄才有几率探寻到矿脉,采集到矿脉可以获得天元矿,每天都可以免费领取30个矿锄,30个领取完之后只能用机缘兑换。同时也可以抢占获得,对非本仙盟的成员进行抢夺获取,并且没有境界压制就算对方境界高也是可以进行抢夺的,尽可能选择比自己开服晚的区会相对
2024-11-23 10:15:14
-
《大江湖之苍龙与白鸟》快速刷级方法
《大江湖之苍龙与白鸟》游戏中玩家的等级可以提升很多的战斗力和能力,但是在升级的过程中还是需要一定的时间的,可能有玩家还不知道怎么升级比较快,下面小编就给大家整理了具体的快速刷级方法,一起来看看吧!《大江湖之苍龙与白鸟》快速刷级方法去襄阳城里和那俩乞丐打架,然后就往树上面退,边退边踩脚趾,空踩脚趾一次
2024-11-23 10:00:12
-
绝区零珂蕾妲贝洛伯格阵容怎么搭配,珂蕾妲贝洛伯格阵容搭配推荐
珂蕾妲贝洛伯格是游戏《绝区零》中的一个角色,该角色不少玩家对其很感兴趣,这里就为大家带来她的最佳阵容推荐攻略,一起来看看吧。绝区零珂蕾妲贝洛伯格最佳阵容搭配推荐珂蕾妲·贝洛伯格+本·比格+11号解析:纯火队,一输出一攻坚一支援,熊哥站场削韧,11号失衡输出最后珂蕾妲连携终结爆炸收场苍角+可琳·威克斯
2024-11-23 09:45:42
-
热血传奇手游:经典三大职业的特点!道士
三、道士职业 道士职业,跟战士职业、法师职业对比,道士职业有可能是相对来说悲惨的一种存在。在热血传奇中道士职业既可以近身作战也可以远攻,而且术士技能多且繁杂,集治愈、召唤、下毒、隐形、控制为一体化,能跟同伴玩出多种多样的配合,可以用剑法与道法,而且还可以用治疗术来帮助别人,不但如此,道士职业还具有惊
2024-11-23 09:30:59
-
仙境传说Ro巴风特之怒:新手超详细入门攻略,助力快速发育!
《仙境传说RO巴风特之怒》手游,是通过端游《仙境传说》复刻的手游,完美继承了端游的游戏特征和乐趣,致力于打造便携、快乐、和谐、的游戏世界。本期就让小编给大家带来这款游戏的新手入门基础攻略教学。角色和职业每位玩家进入游戏时都是“初心者”,在选择职业前都处于这个状态。此时,所有人的起点都是一样的,但可以
2024-11-23 09:16:08